Bacteriophage λ Can Be Modified for Use as a Cloning Vector and Assembled in Vitro
Bacteriophage λ is probably the most extensively studied bacterial virus, and a great deal is known about its molecular biology and genetics. A λ phage virion has a head, which contains the viral DNA genome, and a tail, which functions in infecting E. coli host cells. When λ DNA enters the host-cell cytoplasm following infection, it undergoes either lytic or lysogenic growth. In lytic growth, the viral DNA is replicated and assembled into more than 100 progeny virions in each infected cell, killing the cell in the process and releasing the replicated virions. In lysogenic growth, the viral DNA inserts into the bacterial chromosome, where it is passively replicated along with the host-cell chromosome as the cell grows and divides.
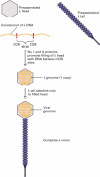
The bacteriophage genome. (a) Electron micrograph of bacteriophage λ virion. The genome is contained within the head. (b) Simplified map of the λ phage genome.
The λ genes encoding the head and tail proteins as well as various proteins involved in the lytic and lysogenic growth pathways are clustered in discrete regions of the ≈50-kb viral genome. When bacteriophage λ is used as a cloning vector, it must be capable of lytic growth, but other viral functions are irrelevant. Consequently, the genes involved in the lysogenic pathway and other viral genes not essential for the lytic pathway are removed from the viral DNA and replaced with the DNA to be cloned. Up to ≈25 kb of foreign DNA can be inserted into the λ genome, resulting in a recombinant DNA that can be packaged in vitro to form virions capable of replicating and forming plaques on E. coli host cells.
During the in vivo assembly of λ virions within infected host cells, viral heads and tails initially are assembled separately, from multiple copies of the various proteins that compose these complex structures. Replication of λ DNA in a host cell generates long multimeric DNA molecules, called concatomers, that consist of multiple copies of the viral genome linked end to end and separated by specific nucleotide sequences called COS sites. Two λ proteins, designated Nu1 and A, bind to COS sites and direct insertion of the DNA lying between two adjacent COS sites into a preassembled head. This process results in the packaging of a single ≈50-kb λ genome from the multimeric concatomer into each preassembled head. Host-cell chromosomal DNA is not inserted into the λ heads because it does not contain any copies of the COS sequence. Once λ DNA is inserted into a preassembled λ head, the preassembled tail is attached, producing a complete virion.

No comments:
Post a Comment